BioUnfold #17 — Where Simulation Now Lives
Simulation has always lived inside biology.
Long before modern AI, it was already doing quiet but decisive work: turning biological understanding into executable systems.
Tumor growth models formalized feedbacks and constraints.
Pharmacokinetic systems made exposure physically accountable.
Transport models expressed how cells and molecules could move.
Chromatin and epigenetic models compressed regulatory logic into mechanistic form.
These systems were never complete descriptions of life.
But they rested on a grammar that had become relatively stable: transcription, translation, degradation, transport, signaling, chromatin regulation, phase behavior, pharmacokinetics.
This grammar is not enough to predict biology.
But it is enough to make biological assumptions executable.
Simulation’s original role was therefore not prediction, but mechanistic bookkeeping. It made stories explicit. It forced levers to be named. It allowed biological reasoning to be decomposed, tested for coherence, and recomposed.
Simulation made biology thinkable.
It did not yet make it explorable.
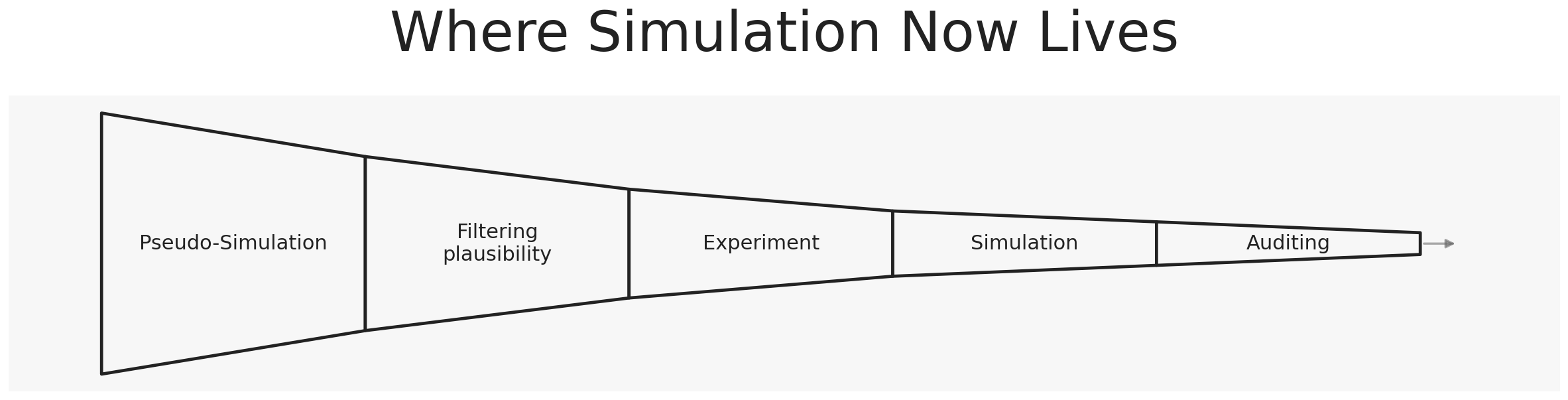
Pseudo-simulation
Machine learning did not introduce a new biological grammar. It turned an existing one into an interactive object.
When sufficient data is available, deep learning turns mechanistic and semi-mechanistic systems into fast, navigable objects — pseudo-simulations built from biological grammar. The burden has shifted: biology is no longer only what simulations try to reproduce. It is increasingly what selects between large spaces of in silico behaviors.
Simulation was long used to ask:
“Can this explanation reproduce what happened?”
It is now increasingly used — often through learned surrogates — to filter:
“What kinds of behavior does this explanation even allow?”
Docking no longer only ranks binders; it constrains which interaction histories are even reachable.
Structure models no longer only describe proteins; they bound how proteins can move under intervention.
PK systems no longer only fit curves; they eliminate drug concepts that cannot exist as treatments.
Simulation moves from validating biology to filtering and structuring biological plausibility.
What is happening is quieter and more consequential than most replacement debates suggest: simulation is being inserted into the hypothesis engine itself.
It is becoming the place where biological explanations are stress-tested before biology is touched.
Auditing
Modern in silico systems are built from partial biological grammars, trained on mixtures of experimental measurements and simulated structure, and pushed beyond the regimes they were explicitly constructed to describe. The futures they generate are therefore not guaranteed to be coherent.
That is why an older role is quietly reasserting itself.
We already see this in places that are publicly visible. Structure prediction is now routinely used to audit mechanistic stories, not just to fill in missing models. Docking and molecular dynamics are increasingly applied to explore interaction regimes and kinetic behaviors rather than to rank binders. Exposure and systems models are used to show when proposed programs cannot exist as treatments at all.
In each case, simulation is not being asked to predict outcomes, but to confront whether the causal objects and dynamics a program relies on are even sufficient.
Condensates are a good example. Many interventions that “affect condensates” do not change abundance, but sequestration. They reshape exchange rates, buffering, and availability. Two compounds can dissolve the same structure and yet act through completely different physical regimes — shifting interaction propensities, or altering compartmentalization dynamics. Those differences often collapse into the same endpoint. But they imply different system trajectories. Simulation is one of the only places where those distinctions have to be made explicit and testable.
Wet experiments are very good at telling us that something moved. They are much worse at forcing biological stories to commit to how a system must evolve once touched.
Simulation does.
It does not aim to close biology.
It asks a lower, more operational question:
Are the causal effects this program is betting on even present in its explanation?
In this sense, in silico systems are becoming a new kind of biological instrument: not one that predicts outcomes, but one that forces mechanisms to take the form medicine will encounter — executable behaviors over time.
Conclusion
Simulation has always lived in biology.
What has changed is not its presence, but its position.
It is no longer only the place where explanations are checked after the fact.
It is becoming the place where biological explanations are forced to become executable before reality is allowed to answer.
In silico systems are not replacing experiments.
They are becoming the only instruments that compel mechanisms to commit to behavior — revealing whether a program’s biological story is even sufficient to support the futures it is trying to build.
And as those futures collide with reality, simulation becomes part of the learning loop itself: where incoherence is not hidden, but used to reshape the models that generate the next set of biological possibilities.